El corazón
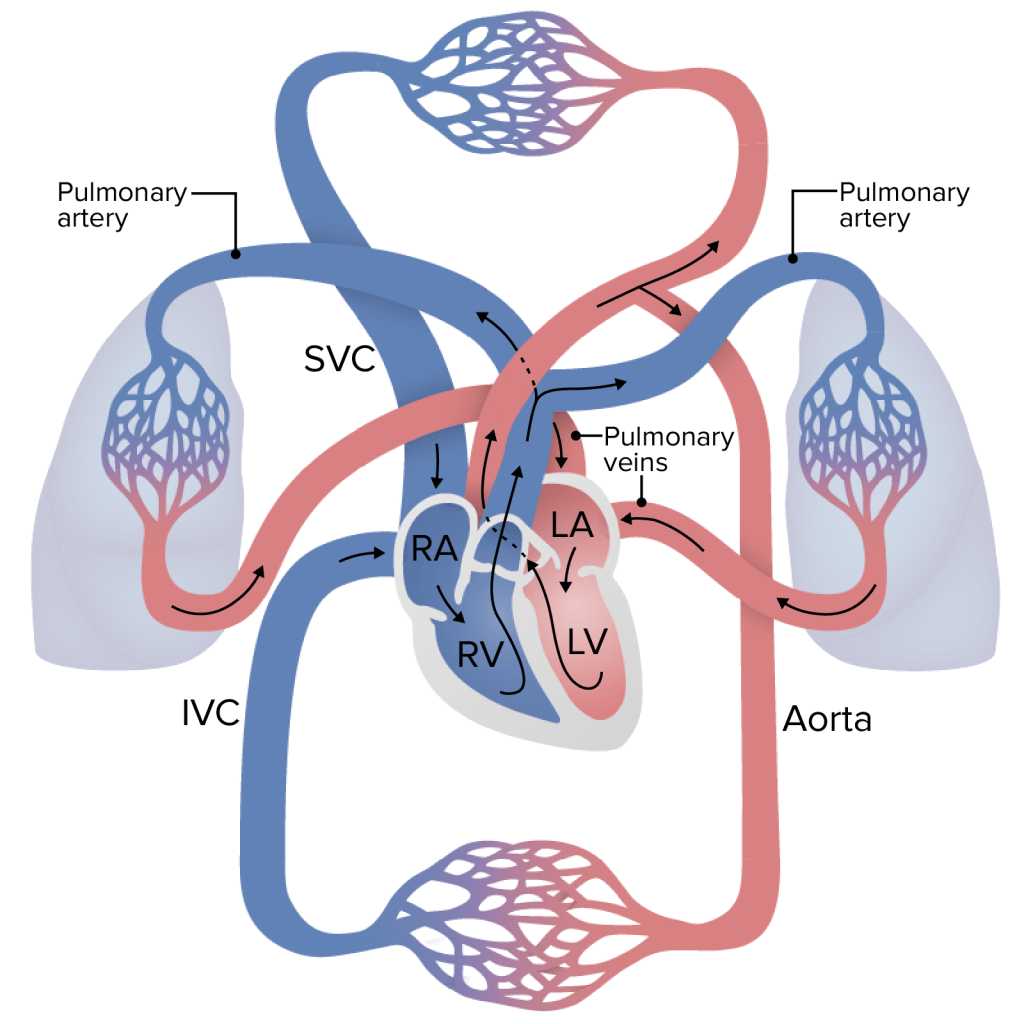
El corazón funciona como un sistema de dos bombas separadas que actúan juntas, el corazón derecho e izquierdo, a su vez, ambas bombas están separadas por las válvulas auriculo ventriculares, en dos cámaras; aurículas y ventrículos. El corazón derecho lleva sangre desde el resto del cuerpo al pulmón desde el ventrículo derecho, mientras que el corazón izquierdo recibe sangre oxigenada desde el pulmón y la reparte por todo el cuerpo, usando la acción de bomba del ventrículo izquierdo.
Existen tres tipos de fibras musculares cardíacas, auricular, ventricular y de conducción. Las auriculares y ventriculares son similares a las fibras de músculo estriado, pero su contracción es más larga, las de conducción eléctrica poseen muy pocas estructuras contráctiles, en cambio de contraen rítmicamente y comandan la actividad eléctrica del corazón. Desde el punto de vista histológico, existen múltiples conexiones entre las fibras, lo que permite el libre flujo de iones a través de sus membranas, haciendo que el corazón funcione como un sincitio, es decir cuando se contrae una fibra el impulso se propaga a las demás. Este sistema también se divide en dos, pero, esta vez, lo hace en sincitio auricular y ventricular, ambos están separados por un anillo fibroso que forma las válvulas aurículoventriculares que no permiten el paso del impulso nervioso. Este sistema hace que las aurículas se contraigan un poco antes que los ventrículos, permitiendo la acción de bomba cardíaca.
Además, el corazón cuenta con un esqueleto fibroso conformado por las válvulas auriculoventriculares, que separan aurículas de ventrículos y las semilunares, aórtica y pulmonar que regulan el paso de sangre hacia sus arterias homónimas. El esqueleto fibroso cumple la función de dirigir la sangre en el sentido correcto y detener el impulso eléctrico proveniente de la red de Purkinje que veremos más adelante.
Potencial de acción del músculo cardíaco
Todas las células del cuerpo poseen un potencial de acción, algunas células son despolarizables debido a la capacidad de intercambiar iones a través de la membrana plasmática. Las células miocárdicas, al igual que el músculo estriado, poseen la capacidad de despolarizarse y contraerse, por estímulos externos o auto despolarizarse como las células del nodo sinusal y sinoauricular.
La relación iónica intra y extracelular del miocardio, dada por la acción de la bomba de protones, que intercambia 3 iones sodio por 2 iones potasio, hace que las membranas estén constantemente polarizadas, aproximadamente a -90 milivoltios, este estado es llamado fase 4 del potencial de acción.
Al recibir un estímulo nervioso, se abren canales rápidos de sodio y este ion ingresa por gradiente químico y eléctrico al medio intracelular, despolarizando la membrana plasmática debido a un aumento brusco de voltaje, que llega hasta +20 milivoltios. Este fenómeno da inicio al período refractario absoluto, momento en el que las células no pueden seguir despolarizándose y, por lo tanto, ningún estímulo será capaz de desencadenar un impulso nervioso. Este momento es llamado fase 0 del potencial de acción y coincide con el inicio de la contracción miocárdica.
Tras el brusco cambio de en la membrana celular se abren canales de potasio regulados por voltaje lo que inicia una salida, por gradiente eléctrico, de este ion, este fenómeno inicia una pequeña repolarización característica de la fase 1 del potencial de acción.
La pequeña repolarización generada por la salida de potasio desde el medio intracelular es rápidamente contrarrestada por la entrada de ion calcio por gradiente de concentración a la célula miocárdica, lo que genera una meseta en el potencial de acción mediada por el efecto antagónico de la salida de potasio y la entrada de calcio. La meseta del potencial de acción coincide con la contracción miocárdica y la duración de esta etapa determina la fuerza de contracción. El miocardio auricular posee una fase de meseta corta lo que determina menor fuerza en la contracción y una deflexión más pequeña en el electrocardiograma, llamada onda p, mientras que las fibras ventriculares poseen una fase de meseta mucho más largas, lo que genera que los ventrículos se contraigan con más fuerza y que la deflexión electrocardiográfica que representa este fenómeno sea mucho mayor, esto da origen a la onda R. Este fenómeno es llamado fase 2 del potencial de acción.
Cuando el gradiente electroquímico de calcio se detiene continua la repolarización iniciada en fase 1, dando fin al período refractario absoluto e inicio a la fase 3 del potencial de acción y al período refractario relativo, debido a la constante repolarización por la salida de potasio al medio extracelular y la caída constante del voltaje transmembrana.
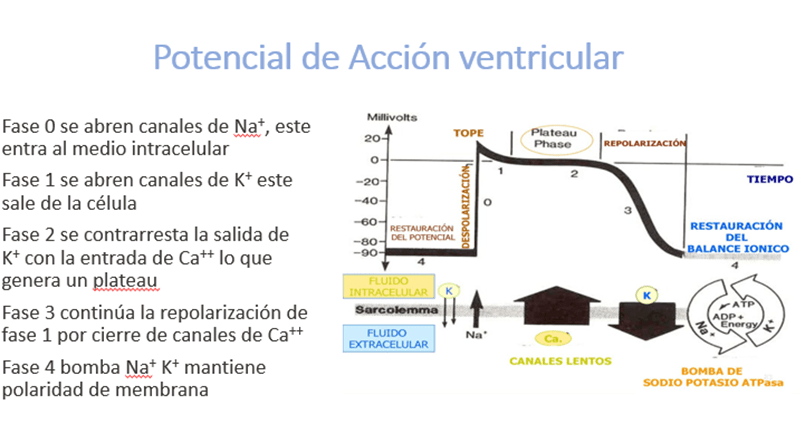
Cuando la célula alcanza el voltaje basal termina el período refractario relativo y la célula vuelve a ser totalmente despolarizable, como ocurre en la fase 4 del potencial de acción. Desde el punto de vista electrocardiográfico la repolarización ventricular genera una deflexión llamada onda T.
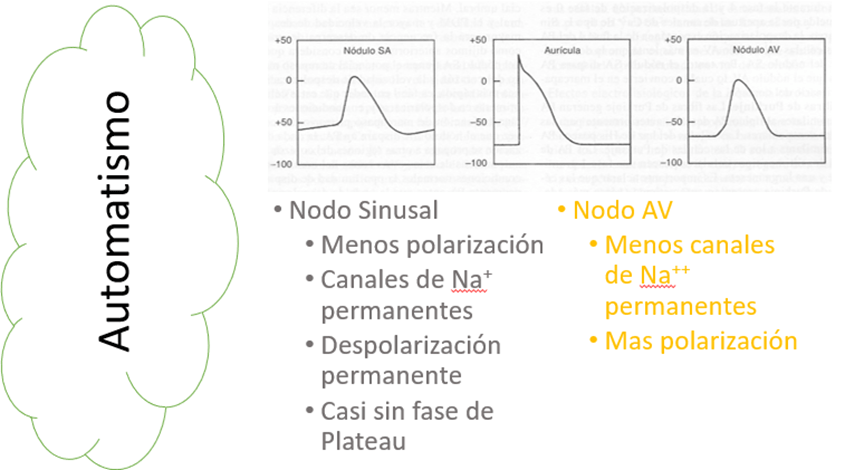
Como se mencionó anteriormente las células miocárdicas son distintas según su función, las del aparato exitoconductor, poseen escasa capacidad contráctil y su potencial de acción ocurre de manera distinta que en el resto del miocardio, ya que poseen canales de sodio constantemente abiertos, lo que implica una polarización menor, su potencial de acción es aproximadamente -60 milivoltios en el nodo sinusal y -70 en el auriculoventricular. La despolarización constante hace que la fase 4 del potencial de acción tenga una pendiente creciente hasta los -50 milivoltios. En ese momento se abren canales de sodio regulados por voltaje y se inicia la fase 0, esto hace que las células del sistema exitoconductor posean un automatismo cíclico. La fase 0 es de pendiente menos pronunciada, que en el miocardio ventricular y auricular, y no posee fase 2 de meseta. El nodo aurículoventricular posee menos canales de sodio permanentemente abiertos, lo que enlentece su automatismo y permite que el nodo sinusal comande la velocidad de contracción cardíaca.
Ciclo cardíaco
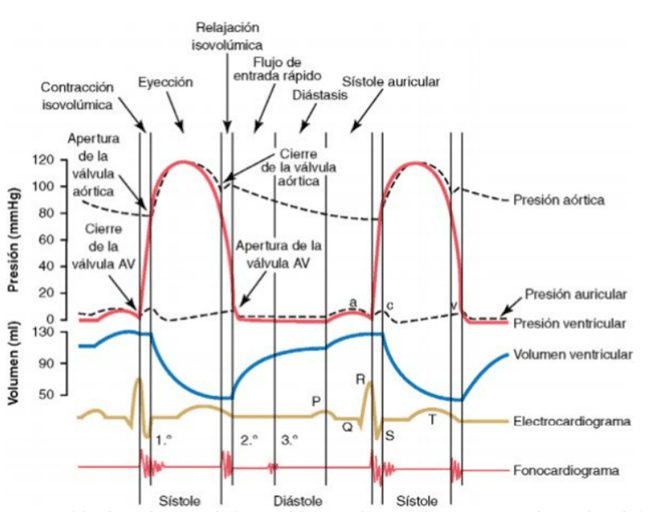
Los fenómenos que ocurren desde el inicio de un latido hasta el comienzo del siguiente son denominados ciclo cardíaco.
El impulso cardíaco nace en el nodo sinusal y viaja hasta el nodo aurículoventricular donde es retrasado unas milésimas de segundo, lo que permite un desfase del sincitio auricular, respecto del ventricular, permitiendo que el primero actúe como bomba de cebado del segundo.
El ciclo cardíaco está formado por un período de relajo llamado diástole, seguido de uno de contracción, llamado sístole. A frecuencias cardíacas normales la diástole corresponde al 35% del ciclo mientras que la sístole equivale al 65%, aproximadamente.
La frecuencia cardíaca determina la duración del ciclo, a mayor frecuencia menor tiempo de ciclo cardíaco, esto no ocurre de manera lineal, así, mientras mayor es la frecuencia cardíaca el tiempo de sístole es menor al 65% normal, por lo tanto, el corazón no alcanza un llenado completo durante el siguiente ciclo cardíaco.
La sangre venosa, proveniente del cuerpo, ingresa a las aurículas desde las venas cavas inferior y superior durante la diástole y, el 80% de esta pasa directamente al ventrículo izquierdo, el restante 20% abandona la aurícula tras su contracción, lo que produce un pequeño aumento de la presión auricular. Con la sangre en el ventrículo comienza la sístole ventricular, la sangre empuja las válvulas auriculoventriculares cerrándolas y los músculos papilares evitan que estas se abran hacia las aurículas, de esta forma se evita el paso de sangre en sentido retrógrado, mientras, las válvulas pulmonar y aórtica permanecen cerradas, lo que aumenta considerablemente la presión ventricular, este corto período de la sístole es llamado contracción isovolumétrica. Cuando los ventrículos tienen la presión suficiente para abrir las válvulas pulmonar y aórtica eso ocurre y la sangre es eyectada con fuerza a la circulación pulmonar y sistémica aumentando la presión en la arteria aorta, una vez que el flujo de sangre aórtico decae ocurre el cierre de su válvula, mientras las auriculoventriculares siguen cerradas, esta parte de la sístole es llamada relajación isovolumétrica y ocurre hasta la apertura de las válvulas auriculoventriculares momento en que comienza la diástole, durante la repolarización ventricular, las aurículas y ventrículos comienzan nuevamente su fase de llenado, iniciando un nuevo ciclo. El cierre de las válvulas aurículoventriculares y semilunares originan los tonos cardíacos
El volumen ventricular al final de la diástole es llamado volumen telediastólico y el residual tras la sístole es llamado telesistólico. El porcentaje del volumen telediastólico que es eyectado durante la sístole, es llamado fracción de eyección y en condiciones normales es aproximadamente un 60%.
Precarga y poscarga
La precarga es la tensión del músculo cuando comienza a contraerse, ese parámetro físico se encuentra determinado por el volumen telediastólico. La poscarga es la resistencia contra la cual el corazón debe empujar la sangre, en términos prácticos la poscarga es la resistencia que ejerce la aorta al flujo proveniente del ventrículo izquierdo.
Sistema de excitación cardíaca
La excitación y conducción eléctrica cardiaca ocurre gracias a un grupo de células musculares especializado en estas tareas. Su función es entregar ritmo y coordinación a la acción de bomba del corazón.
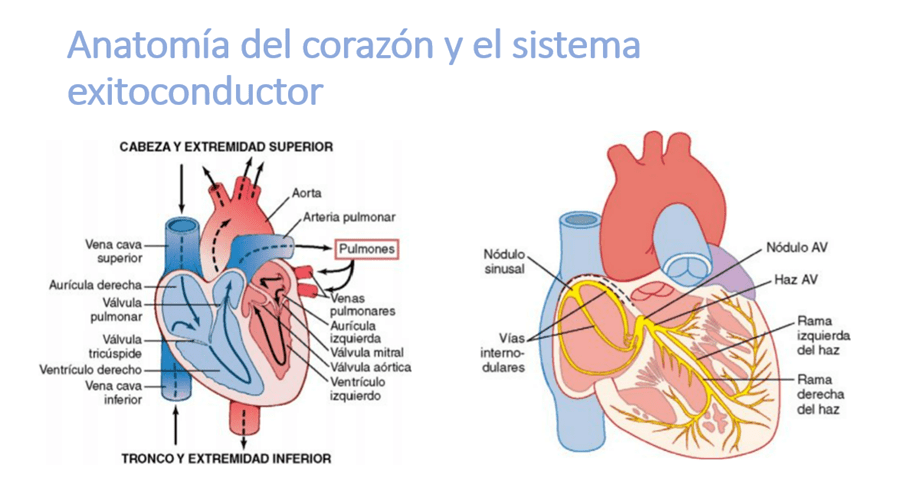
En condiciones normales el estímulo nace desde el nodo sinusal, que está situado cercano a la desembocadura de la vena cava superior y la salida de la arteria aorta, posee conexiones directas con el resto de la musculatura auricular, por eso, todos los impulsos que nacen desde el nodo sinusal se propagan directamente a las aurículas.
El impulso nervioso viaja a través del miocardio auricular y por las vías internodulares hacia el nodo auriculoventricular, que se encuentra posterior a la válvula tricúspide, en el se retrasa el impulso nervioso para permitir la coordinación entre aurículas y ventrículos, esto favorece el llenado ventricular previo a la fase de eyección. El retraso se debe a un aumento de la resistencia al flujo de iones debido a una disminución de uniones celulares.
Una característica importante del nodo aurículoventricular es la unidireccionalidad, que condiciona la imposibilidad de transmitir impulsos eléctricos desde el ventrículo a la aurícula, evitando la reentrada de impulsos nerviosos
El impulso retardado en el nodo auriculoventricular, pasa al haz auriculoventricular, que se divide en una rama izquierda y otra derecha, para, finalmente ramificarse en la red de Purkinje. La red de Purkinje está formada por células incluso más grandes que las ventriculares y su velocidad de conducción es aproximadamente 6 veces más que la de los miocitos ventriculares, lo que permite una transmisión instantánea del impulso nervioso a los ventrículos.
Regulación de la contractilidad cardíaca
El corazón es capaz de latir por si solo gracias al automatismo de despolarización de las células de su sistema exitoconductor, descrito con anterioridad. Los mecanismos capaces de regular el automatismo son intrínsecos y extrínsecos. De esta forma el sistema cardiovascular es capaz de regular el gasto cardíaco, que se entiende como la frecuencia cardíaca multiplicada por el volumen de eyección. El gasto cardíaco normal es entre 4 a 6 L/min, pudiendo duplicarse en condición de ejercicio o enfermedad.
Gran parte de la sangre que bombea el corazón está determinada por el retorno venoso, este fenómeno es llamado ley de Frank-Starling, esta versa que mientras más sangre ingresa por retorno venoso, mayor será el gasto cardíaco. Esto ocurre por tención de las fibras miocárdicas, a mayor presión intracardiaca mayor velocidad y fuerza de contracción, por lo tanto, mayor gasto cardíaco.
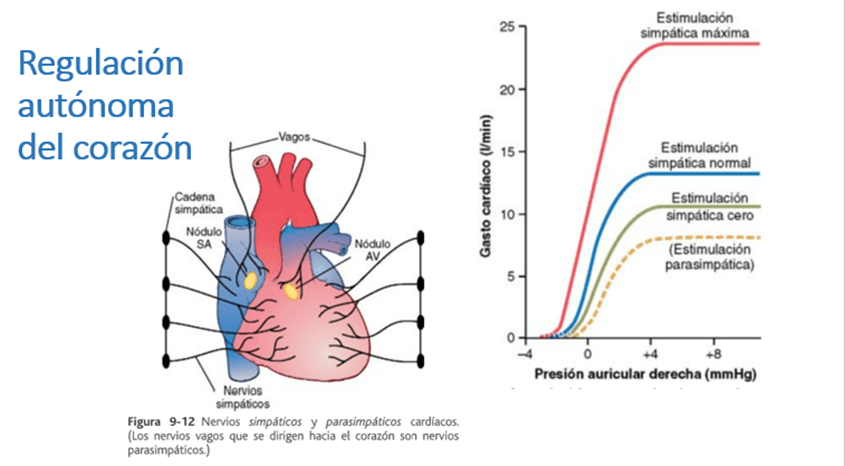
Otro sistema directamente relacionado con la regulación del gasto cardiaco es el sistema nervioso autónomo, bajo estimulación simpática aumenta la contractilidad y frecuencia, mientras que la parasimpática la disminuye.
Los nervios vagos están distribuidos por todo el corazón, actúan a través de la acetilcolina, que abre canales de potasio, hiperpolariza la célula y enlentece el estímulo en los nodos sinusal y auriculoventricular, que están ampliamente inervados por sus fibras.
Los nervios simpáticos se encuentran ampliamente relacionados con la inervación de la musculatura ventricular y del resto de las estructuras cardíacas, su neurotransmisor es la noradrenalina, que actúa abriendo canales de calcio y sodio, lo que despolariza al miocardio y aumenta su frecuencia y fuerza contráctil.
Como se mencionó anteriormente los iones potasio y calcio, juega un rol importante en la regulación de la fuerza contráctil. El rol de ion potasio en la regulación del del ciclo cardiaco es repolarizar la membrana miocárdica cuando está despolarizada. Cuando la célula se encuentra polarizada el exceso de potasio extracelular despolariza parcialmente la célula. por lo tanto, un exceso de potasio tiende a producir bradicardia y debilitar la contracción muscular. El exceso de iones calcio genera lo contrario a la hiperpotasemia, sin embargo, la regulación de este ion está estrechamente controlados, por lo que los cambios de las concentraciones de calcio rara vez tiene implicancias clínicas.
La temperatura es otro factor que regula el gasto cardíaco, en presencia de calor o fiebre la frecuencia cardiaca aumente por aumento en la permeabilidad de membrana a iones, lo que aumenta el automatismo del corazón. Durante la hipotermia ocurre lo contrario.
Otro mecanismo que puede regular el automatismo es la presión arterial, se ha observado que presiones medias sobre 160 mmHg pueden disminuir el gasto cardíaco.
Circulación
La circulación sanguínea cumple variadas funciones necesarias para la vida, dentro de ellas encontramos el transporte de nutrientes, detritus y hormonas, para mantener condiciones adecuadas destinadas a un correcto funcionamiento del organismo.
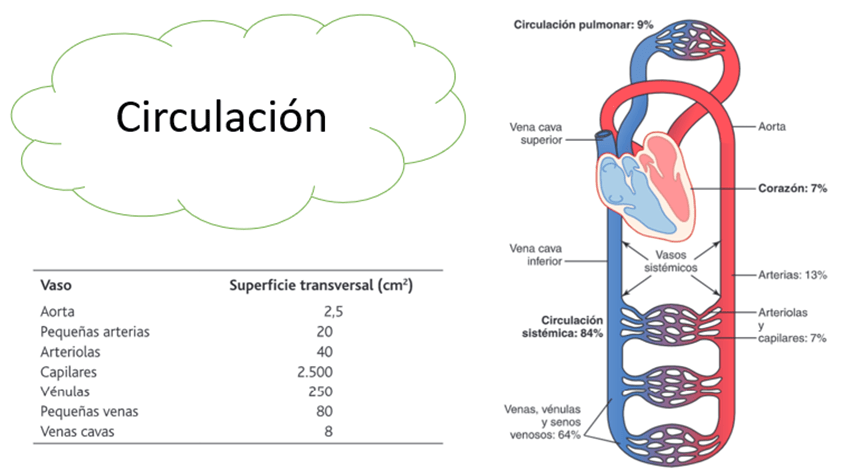
Para satisfacer las funciones vitales, la circulación se divide en dos; sistémica y pulmonar. Ambas a su vez están formadas por una serie de vasos sanguíneos con distintas características, llamados arterias, arteriolas, capilares, vénulas y venas.
Las arterias son rígidas y transportan la sangre con una presión y velocidad alta hacia los tejidos. Las arteriolas conforman el punto final de las arterias y actúan como llave de paso para una liberación de sangre controlada hacia los capilares, esto gracias a su fuerte pared muscular. El intercambio de sustancias y detritus ocurre en los capilares, poseen paredes muy delgadas y tienen abundantes poros permeables al agua y moléculas pequeñas. La sangre de los capilares fluye hacia las vénulas que aumentan progresivamente su diámetro hasta convertirse en venas, que son muy distensibles, por lo que funcionan como reservorio sanguíneo, y transportan sangre desde las vénulas hasta el corazón. Las venas poseen una pared delgada, pero muscular, que permite contraerse y así regular el retorno venoso en condiciones de ejercicio, o dilatarse, y reservar sangre, en condiciones de reposo.
El 84% de la sangre del cuerpo se encuentra en la circulación sistémica, mientras que el 16% está en la circulación pulmonar. El 64% de la circulación sistémica se encuentra en las venas, esto debido a que la distensibilidad venosa es 8 veces la arterial, lo que hace que a un mismo diámetro las venas puedan acumular 8 veces la sangre que acumulan las arterias, casi sin cambios de presión. Es sorprendente que solo 7% de la sangre se encuentra en los capilares y otro 7% ocupa el corazón.
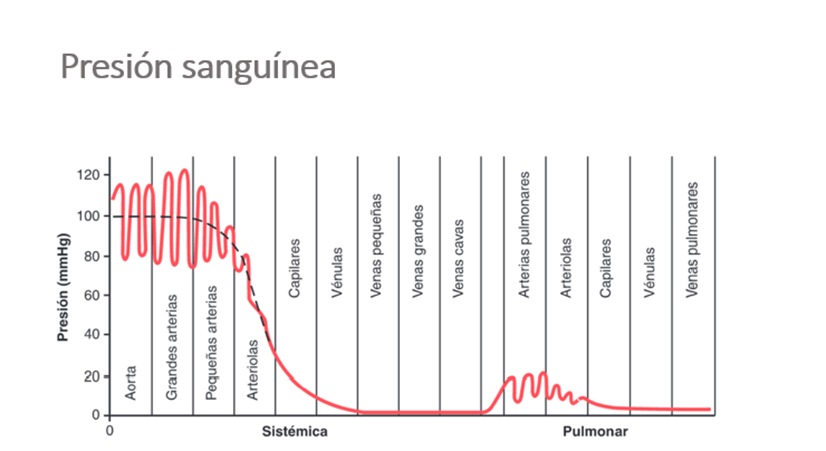
La presión sanguínea es decreciente a lo largo del sistema vascular, en la aorta oscila los 100 mmHg, a este nivel es pulsátil, la sistólica puede llegar hasta los 120 mmHg y la diastólica a los 80 mmHg, en condiciones normales. A medida que se profundiza el sistema arterial aumenta su diámetro transversal, lo que produce una brusca disminución de la presión, así, en las arteriolas puede llegar a 35 mmHg, en los capilares a los 17 mmHg, presión suficiente para permitir el intercambio de plasma y moléculas pequeñas, finalmente, la presión en las venas cavas es 0.
En la circulación pulmonar la presión es menor, pero tienen un comportamiento similar. Las arterias pulmonares son pulsátiles y su presión promedio es de 25 mmHg la sistólica y 8 mmHg la diastólica.
La distensibilidad de los vasos sanguíneos es un componente importante del sistema circulatorio y determina en muchos casos la presión arterial. Cuando ocurre una disminución o incremento brusco del volumen sanguíneo los vasos responden cambiando su diámetro gracias a su musculatura lisa, este fenómeno es llamado distensibilidad diferida. Cuando se incrementa el volumen sanguíneo bruscamente, la presión arterial aumenta en la misma proporción, sin embargo, al cabo de unos minutos el vaso sanguíneo se distiende diferidamente y la presión arterial vuelve a caer por relajación de su musculatura lisa, fenómeno conocido como “relajación por estrés”. Este fenómeno permite que la sangre pueda fluir durante la diástole y llegue a tener flujos laminares en los capilares.
Principios básicos de la función circulatoria
- La velocidad del flujo sanguíneo se controla en relación a la necesidad del tejido. Cuando un tejido requiere mayor aporte de nutrientes, el déficit de estos o el exceso de detritus es detectado por la circulación local, lo que regula el paso arteriolar de sangre hacia los tejidos, ajustando el flujo a sus demandas.
- El gasto cardíaco se controla principalmente por la suma de todos los flujos tisulares locales. Cuando muchos tejidos requieren mayor volumen de sangre aumenta el retorno venoso y esto aumenta el gasto cardíaco.
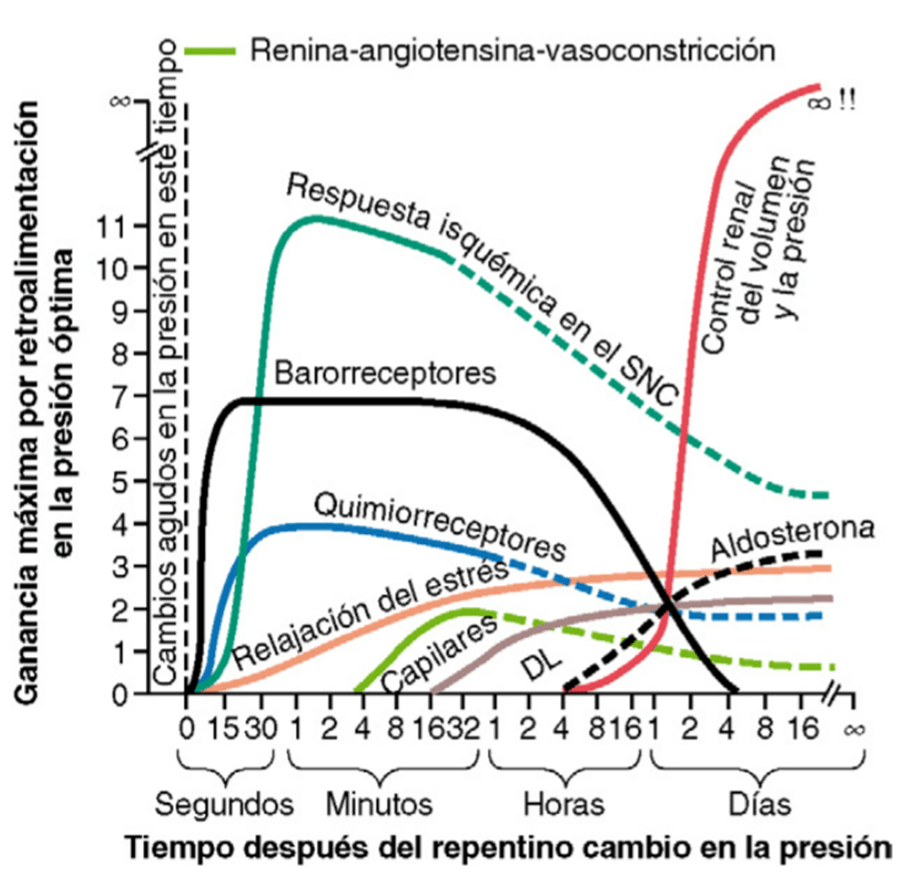
- La regulación de la presión arterial es independiente del control del flujo sanguíneo local o del control del gasto cardíaco. La presión arterial está fuertemente regulada a nivel central y ante una brusca caída la respuesta se inicia en segundos y se puede prologar por días cuando se agregan factores renales de control de la presión. Esta respuesta es independiente de las necesidades fisiológicas de los tejidos.
Mecanismos de control del flujo sanguíneo
El control del flujo sanguíneo local está fuertemente regulado por la necesidad de nutrientes y hormonas y la eliminación de desechos, como dióxido de carbono e hidrogeniones a nivel local.
Esas necesidades hacen que los órganos encargados de la depuración de la sangre o las glándulas secretoras de hormonas se encuentren muy irrigadas, como ocurre con la glándula tiroides, el riñón o el hígado. Mientras que otros sistemas que en estado de reposo reciben escaso flujo sanguíneo, a pesar de constituir gran masa corporal, como el tejido muscular.
El control del flujo sanguíneo local puede dividirse en dos fases, control a corto plazo o largo plazo.
- El control a corto plazo, regula el diámetro vascular, a través de vasoconstricción o vasodilatación, puede ocurrir en segundos o minutos.
La saturación arterial de oxígeno es un factor importante de control a corto plazo, en ausencia de oxígeno el flujo sanguíneo puede llegar a cuadruplicarse. Otros nutrientes también han demostrado tener efectos en la vasculatura, por ejemplo, la falta de glucosa tisular puede generar vasodilatación.
- El control a largo plazo ocurre en un lapso de tiempo de horas, días, semanas o incluso, meses, son más precisos que los de corto plazo y ocurren como consecuencia de cambios en el tamaño corporal y en el número de vasos sanguíneos que nutren un determinado tejido.
Cuando los cambios en la presión arterial se mantienen en el tiempo, el sistema vascular responde llevando el flujo de sangre a los tejidos casi a niveles normales, esto puede ocurrir cuando una persona sufre de hipertensión arterial o cuando las demandas metabólicas de un tejido cambian brusca y persistentemente en el tiempo, por ejemplo, cuando comienza a vivir en altura o pasa de una condición sedentaria a otra activa.
Algunas respuestas de largo plazo son; la neovascularización, el desarrollo de circulación colateral.
Control en territorios específicos
- Riñones: Cuando se filtra demasiado líquido de la sangre a través del glomérulo hacia el sistema tubular, las señales de retroalimentación de la mácula densa generan vasoconstricción de las arteriolas eferentes, disminuyendo la diuresis.
- En el cerebro la concentración de dióxido de carbono y de hidrogeniones es de gran importancia, el aumento de cualquiera de ellos genera vasodilatación cerebral y un rápido lavado. Este estricto mecanismo responde a la especial sensibilidad del tejido cerebral a los cambios de pH.
- En la piel los cambios de diámetro vascular responden fuertemente a cambios de temperatura, en presencia de calor el flujo sanguíneo de la piel puede llegar a triplicarse, mientras que, en temperaturas bajas, la irrigación de la piel puede ser cercana a 0.
Algunas sustancias son secretadas directamente por el endotelio vascular y poseen un rol en el control local del flujo sanguíneo, las dos más importantes son el óxido nítrico, potente vasodilatador liberado por células endoteliales sanas, y la endotelina, liberada por endotelio enfermo, que actúa como vasoconstrictor.
Control humoral de la circulación
El control humoral de la circulación se refiere al control por sustancias segregadas o absorbidas en los líquidos del organismo.